m6A研究又有新手段了?赶紧来了解一下吧!栏目:研究动态发布时间:2019-08-14RNA甲基化m6A是如今的研究热点之一,Cell上新发表了一篇介绍不使用m6A抗体的检测mRNAm6A水平的Resource文章......RNA甲基化m6A是如今的研究热点之一,目前主要的研究思路包括差异表达的write,reader和eraser基因分析;m6A抗体检测全转录组甲基化水平;分析m6A甲基化水平变化的靶基因和下游机制研究。而在7月25日的Cell上新发表了一篇介绍不使用m6A抗体的检测mRNAm6A水平的Resource文章,小编时间和大家分享一下。作者开发这一方法的前提是有研究报道了MazF能特异性的识别无修饰的ACA序列并发挥RNA酶活性在ACA之前剪切底物。但ACA序列的个A发生m6A甲基化之后将无法被MazF所识别,如图一所示。图1MazFRNA酶作用示意图根据这一原理,作者将纯化后的mRNA进行MazF酶处理,然后再对打断的RNA进行逆转建库测序。对于无修饰的位点,ACA处被完全剪切,测序reads正好分布于该位点上下游而且比对至该处的上下游reads数是相同的.如图2所示。而甲基化位点的reads会包含上下游的序列。作者开发了MAZTER-MINE软件包专门进行分析(图3)。分享的内容能对大家有所帮助,想要了解更多,欢迎致电咨询。湖南科研技术服务实验
转录组测序结果及TCGA数据库分析)图5RNA-Seq和m6A-seq联合鉴定SOCS2是介导的m6A修饰的下游靶基因PLoSOne2015,在许多不同种类的RNA中,都已观察到N6-腺苷(m6A)的甲基化,但其在microRNAs中还没有被研究。研究者在FTO1C1,FTO2D4和FTO3C3细胞系中,通过敲除m6A甲基转移酶FTO筛选到表达差异的microRNA,说明miRNA受m6A甲基化的调控。进一步通过MeRIP-Seq发现相当一部分的microRNA具有m6A修饰。通过motif分析,他们发现了区分甲基化和非甲基化microRNA的一致序列。该文章所述的表观遗传修饰在基因表达的转录调控的复杂性上增加了一个新的层次。图FTO敲除对甲基化的miRNAs的稳定状态的影响。参考文献Y,DominissiniD,RechaviG,HeC:Geneexpressionregulationmediatedthroughreversiblem(6)(5):'(1):(12):(6):(1):(1):(uridine)(41):(6)(7540):(1):(7481):(4):(6)A-LAIC-seqrevealsthecensusandcomplexityofthem(6)(8):'UTRm(6)(4):(7544):(6)(6):(5):(7667):(2):"">panstyle="color:#f5c81c;">xiainducesthebreastcancerstemcellphenotypebyHIF-dependentandALKBH5-mediatedm(6)(14):"">panstyle="color:#f5c81c;">(40):(6)(3):(1):(1):(4):(11):。湖南兔科研技术服务购买动物疾病模型还为科研人员提供了研究人类疾病的跨学科方法。
RNA各种可逆的化学修饰被认为是一种新的表观遗传调控方式。m6A是真核生物mRNA常见的化学修饰,在调控mRNA稳定性,剪切和翻译方面具有重要的作用。作者使用转录组测序发现了METTL3(甲基转移酶3),一种主要的RNAN6-腺苷-甲基转移酶,在人肝细胞(HCC)和多种实体中高表达。在临床上,METTL3的过度表达与肝细胞患者不良预有关。体外实验证明敲除METTL3会抑制HCC细胞增殖,迁移及克隆形成。体内实验证明敲除METTL3会明显抑制HCC体内成瘤和肺转移。另外,使用CRISPR/dCas9-VP64系统,内源性高表达METTL3会促进HCC细胞在体外和体内生长。通过转录组测序、m6A-Seq、MeRIP-PCR,作者确定了SOCS2(细胞因子信号2的抑制因子)作为METTL3介导的m6A修饰的下游靶基因。敲除METTL3表达会消除SOCS2mRNAm6A修饰并增强SOCS2mRNA表达。m6A介导的SOCS2mRNA降解是依赖于m6A“读取器”蛋白YTHDF2。总之,METTL3在HCC大部分高表达中,并通过m6A-YTHDF2依赖机制抑制SOCS2表达从而促进HCC进展。因此,作者发现了在肝发生过程中表观遗传改变的一种新机制。图4RNA甲基化转移酶METLLT3在肝组织中高表达。
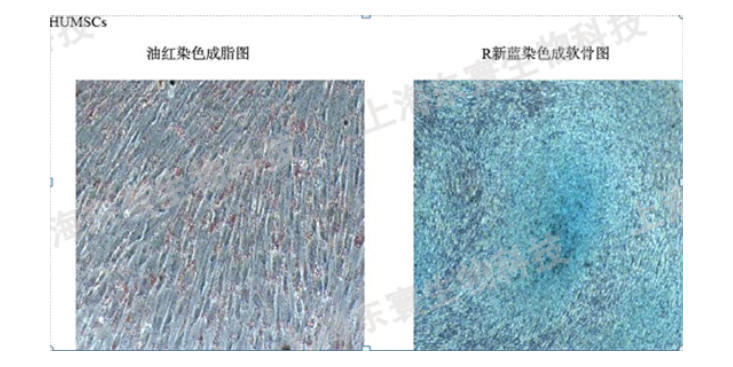
外泌体的功能外泌体可能作为细胞之间物质和信号通讯的途径,不同的细胞通过分泌携带不同组分的外泌体实现细胞间通讯,这些外泌体被受体细胞吸收,通过物质交换或释放内含物实现物质和信号的交流。外泌体与受体细胞的信息交流可能通过以下4种途径实现:外泌体表面膜蛋白质被目的细胞表面的受体识别,从而靶细胞;外泌体在蛋白酶作用下产生的表面膜蛋白质碎片可作为目的细胞表面受体的配体;外泌体脂膜可以与目的细胞膜融合,将蛋白质和RNA等内容物释放进去,此类膜融合可能改变靶细胞的一些膜特性,例如脂质和膜蛋白质的密度及种类;目的细胞通过胞吞的形式摄入外泌体经由以上几种途径,外泌体将其携带的生物信息运输到周边靶细胞或经血液等体液运输而被远处组织细胞摄取,进而影响目的细胞的基本功能和基因表达。几乎所有的细胞都可以在自发或在一定刺激条件下产生外泌体,不同的细胞产生的外泌体具有不同的功能,这些外泌体参与了一系列生理和病理过程,如发生与发展、抗原呈递、免疫调节、组织愈合等。此外,外泌体与疾病的研究表明:外泌体可能在代谢性疾病的出现以及心血管健康中发挥作用。人工模型是指通过人工手段制造的疾病模型。
动物模型在科研中有着普遍的应用。首先,它们可以帮助科研人员深入理解的共同性,即不同物种之间存在的共有理变化过程。通过对动物模型的研究,科研人员可以更清楚地了解的发展过程和机制,为人类的检查提供理论依据。其次,动物模型还为新研发和苗测试提供了的平台。在研发过程中,科研人员可以通过对动物模型进行处理,观察其和副作用,为新的临床试验提供依据。而在苗测试中,动物模型则可以用来评估苗的性和安全性。此外,动物模型还为科研人员提供了研究人类的跨学科方法。例如,通过比较人类和动物模型的基因组学、蛋白质组学等数据,可以发现与发生相关的关键基因和蛋白质,从而为的和检查提供新的思路。虽然动物模型在科研中发挥了巨大的作用,但也存在一些挑战。首先,由于物种差异的存在,动物模型的表现与人类可能存在差异,因此需要谨慎使用。此外,动物模型的伦理问题也不容忽视,科研人员需要在符合伦理规定的前提下进行相关研究。尽管存在挑战,动物模型的发展前景仍然值得期待。随着科技的不断进步,科研人员将能够开发出更为精确、实用的动物模型。动物疾病模型可以分为两种类型:自然模型和人工模型。江苏大鼠科研技术服务实验室
细胞核是细胞的控制中心,它包含了遗传物质DNA,控制着细胞的生长和分裂。湖南科研技术服务实验
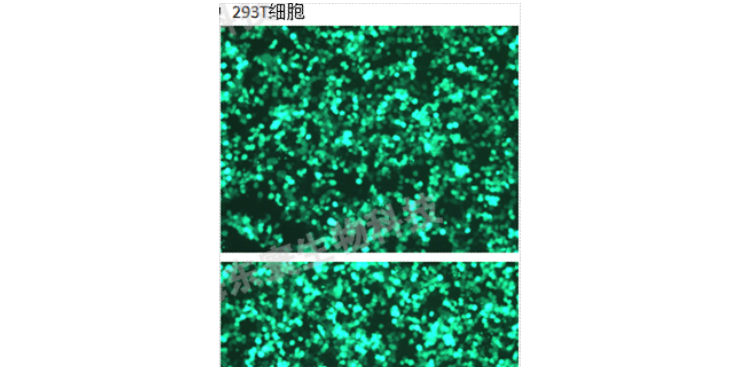
采用opti-MEM和Lipo3000分别转染含有目的基因的pMSCV-eGFP、VSV、GAG质粒及对照载体,每皿加入脂质体-质粒转染混悬液按购买脂质体相关说明书操作定量。继续培养24h。2)24小时后,将培养基更换为新鲜的DMEM完全培养基,放进细胞培养箱继续培养48~72h。3)48~72h后收集上层培养液,并过μm滤膜,采用ELISA法对所获得的慢载体进行滴度测定。如不及时使用可以冻存于-80℃。3、慢转染1)转染前1天将细胞接种6孔培养板,时细胞的融合率约为50%,前需换液,加入1mLDMEM完全培养基。2)冰浴融化后加入相应体积的液及聚凝胺(Polybrene),混匀后放入37℃孵箱中继续培养3)4h后补充1mL培养基,14h后换液(24h内换液即可)。4)72h后用倒置显微镜观察荧光,监测效率,出现较多荧光时将等量的转染细胞和未转染细胞分别加入等浓度Puromycin(Puromycin或其他筛选浓度需要事先摸索)。5)待未转染细胞全部死亡并且可观察到满意荧光量时,降低Puromycin浓度培养。也可以挑去单克隆细胞株进行进一步培养,以得到满意的稳定表达目的基因的细胞株。6)使用qRT-PCR和Westernblot的方法检测目的基因的表达量和蛋白水平是否显著提高。7)由此可得三组细胞株:a.正常细胞株;b.空载载体的细胞株。湖南科研技术服务实验